Tác giả: Tndcong
-

Các loại đảo
Trong nội dung Tiến hóa và Sinh thái của chương trình Sinh học 12, đảo đại dương và đảo lục địa là những khái niệm được nhắc đến nhiều. Vậy đảo là gì? Có những loại đảo nào?
Đảo là phần đất bao quanh bởi nước biển. Tương tự với cấu trúc này là cù lao, dành gọi những phần đất được bao quanh bởi dòng sông. Có nhiều loại đảo khác nhau như: đảo đại dương, đảo lục địa, đảo núi lửa v.v…
Đảo đại dương là đảo được hình thành giữa đại dương. Những hòn đảo loại này luôn luôn nằm ở đỉnh các ngọn núi cao được nổi lên từ đáy đại dương.
Đảo lục địa là phần đất tách ra từ một lục địa rộng nào đó, thường ngăn cách với lục địa bổi một vùng biển. Vì vậy, chúng thường nằm ở giữa biển (chứ không phải đại dương). Điều này xảy ra khi một phần lớn đất chìm xuống hoặc khi mực nước biển dâng lên.
Đảo núi lửa là những hòn đảo được hình thành do những dòng dung nham núi lửa phun trào, chảy xuống biển, nguội lại, hình thành nên. Chúng thường ở gần bờ; trường hợp đặc biệt được hình thành từ miệng núi lửa hãy khe nứt dưới đáy đại dương.
Các đảo lớn nhất trên thế giới theo thứ tự giảm dần về kích thước là: Greenland, New Guinea, Borneo, Madagascar, Baffin, Sumatra, Honshu và Vương quốc Anh. Theo nguồn gốc, các đảo này đều là đảo lục địa hoặc đảo đại dương (không phải được hình thành từ núi lửa).

Đảo Greenland (Canada) 
Đảo New Guinea (Indonesia + Papua New Guinea) 
Đảo Borneo (Malaysia + Brunei) 
Đảo Baffin (Canada) Đảo Honshu (Nhật) Vương quốc Anh Khi loại bỏ phần nước đi, thì các hòn đảo cũng trông như những ngọn núi nhô lên từ dưới đáy đại dương hoặc biển, mà trong hệ động thực vật ở ven bờ, kéo dài ra biển/đại dương giống như những cánh rừng trải dài trên triền núi. Cho đến nay, con người vẫn chưa đến được tận cùng vực thẳm của đáy đại dương. Khu vực sâu nhất mà con người xuống đến và quan sát được cho đến lúc này là khu vực bình nguyên dưới đáy đái dương, sâu vài nghìn mét. Nhưng dưới đó vẫn lại là … một vực đại dương khác sâu trong lòng nước mà các nhà thám hiểm vẫn chưa đi đến được. Nói cách khác, nhìn một cách tổng quan, nếu đi từ thấp lên cao, thì bề mặt trái đất cứ có mặt phẳng, rồi lại đến núi, ngang với nó là vùng bình nguyên rộng lớn; sau đó lại là núi và bình nguyên trong lòng nước biển (đảo và các đại lục); sau đó lại là núi và bình nguyên trên cạn mà chúng ta thường nhìn thấy.
-
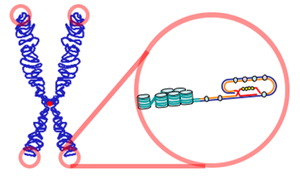
Telomere
Nhân dịp làm semina môn Sinh học Phân tử, người viết có phụ trách nội dung về telomere – một mảng khá “mới” trong chương trình phổ thông, cũng như trong nội dung nghiên cứu khoa học ở Việt Nam. Đây là nội dung không khó, nhưng lại khó để tưởng tượng nên nhiều người khi nghe vẫn không hiểu được. Thiết nghĩ đây là nội dung bổ sung tốt cho những bài học di truyền phân tử trong Sinh học THPT, nên người viết chia sẻ với người đọc nội dung thú vị này. Nội dung này đã được chỉnh sửa cho phù hợp với nhu cầu tham khảo của học sinh THPT.
I. TELOMERE:
1. Telomere là gì?
Trong cơ chế tái bản, DNA được tổng hợp theo hai mạch song song nhau. Sự tháo xoắn được thực hiện từ đầu nọ đến đầu kia của DNA với sự xúc tác của enzyme helicase. Theo tinh thần tiết kiệm và tranh thủ phổ biến trong tất cả các hoạt động sinh lí của cơ thể, mạch kép của DNA tháo xoắn đến đâu, sẽ được các phức hệ enzyme DNA – polymerase I, DNA – polymerase II, DNA – polymerase III và các enzyme khác xúc tác tổng hợp mạch mới ngay đến đó. Vấn đề xuất hiện ở đây là, trong cả 3 hệ enzyme trên đều không có đầu 3’-OH tự do là nơi để các nucleotide tiếp theo liên kết vào, một điều kiện tối cần thiết để chúng có thể thực hiện quá trình tổng hợp kéo dài mạch. May thay, trong tế bào lại có một hệ enzyme khác, RNA – polymerase, tự nó có đầu 3’-OH tự do và do đó có khả năng khởi đầu quá trình tổng hợp kéo dài mạch. Vì thế, cơ chế tự nhân đôi trong nhân tế bào đã “mượn tạm” hệ enzyme này thực hiện quá trình tổng hợp ra đoạn nhỏ mạch RNA (các primer – RNA mồi) bổ sung với mạch khuôn DNA, nhằm tạo đầu 3’-OH tự do giúp DNA – polymerase thực hiện chức năng của mình. Tuy nhiên, việc khắc phục “nhược điểm” này lại để lộ ra một “nhược điểm” khác trong cơ chế tái sinh ở cấp độ phân tử – sự tái bản DNA.
Do mạch DNA chỉ có thể được tổng hợp theo chiều từ 5’ đến 3’ (dưới tác dụng của DNA – polymerase khi trượt trên mạch khuôn theo chiều 3’ – 5’), nên sự tái bản trên hai mạch đơn của DNA diễn ra không giống nhau:
- một mạch được tổng hợp liên tục cùng chiều với sự tháo xoắn DNA gọi là mạch dẫn đầu (leading strand) với chỉ một đoạn primer cho mỗi vị trí khởi sự tái bản (điểm ori);
- một mạch được tổng hợp gián đoạn ngược chiều với sự tháo xoắn DNA gọi là mạch ra chậm (lagging strand hay mạch Okazaki) với nhiều đoạn primer khởi đầu cho từng phân đoạn Okazaki có chiều dài khoảng 1000 – 2000 cặp nucleotide. Các primer này dài từ hàng chục đến hàng trăm nucleotide. Trên mạch ra chậm, sau khi hoàn tất quá trình tổng hợp các phân đoạn Okazaki, DNA – polymerase I với hoạt tính exonuclease sẽ vừa phân giải các primer, vừa tổng hợp đoạn DNA thay thế dựa trên đầu 3′-OH của đoạn DNA đã tổng hợp trước đó.
Như vậy, sau khi kết thúc quá trình tổng hợp kéo dài, tận cùng của DNA ở hai đầu 3’ (của mạch DNA cũ) vẫn còn tồn tại một primer chưa được thay thế. Và các primer này sẽ được các enzyme loại bỏ mà không được thay thế do sự khiếm khuyết của hệ enzyme DNA – polymerase khi không có khả năng tạo đầu 3’-OH trong phức hệ. Chính điều này làm cho hai đầu của DNA bị ngắn đi một đoạn tương ứng với chiều dài của primer (vài chục đến hàng trăm nuceotide) sau mỗi lần nó tự nhân đôi (tương ứng với sự phân chia tế bào). Vậy là, hệ gene của tế bào sẽ bị “thất thoát” sau mỗi lần tạo ra thế hệ mới. Tế bào đã làm cách nào để khắc phục sai sót này?
Các nghiên cứu đã chỉ ra rằng, trong DNA tồn tại những đoạn trình tự lặp lại có chiều dài nhất định ở hai đầu “dự trữ” cho sự mất mát này, nhằm đảm bảo an toàn cho những gene cấu trúc nằm bên trong. Đây chính là các telomere.
Telomere là những trình tự lặp lại của DNA ở các đầu mút của nhiễm sắc thể. Mặc dù cũng được cấu thành từ các đơn phân nucleotide, nhưng telomere không mã hóa cho protein. Ngoài chức năng trên, chúng còn bảo vệ các nhiễm sắc thể trong quá trình phân bào, giữ cho các nhiễm sắc thể không dính vào nhau.
2. Lịch sử tìm thấy telomere:
3. Cấu tạo telomere:
Telomere được tìm thấy trong hầu hết các tế bào nhân thực và một vài tế bào loài sinh vật nhân sơ.
Sinh vật nhân thực (Eukaryote) có nhiễm sắc thể dạng mạch thẳng, và vì vậy mà gặp phải những vấn đề trong quá trình tái bản ở hai đầu mút mỗi nhiễm sắc thể như đã nói ở trên. Sinh vật nhân sơ (Prokaryote) có DNA dạng vòng, nên không có các đầu tận cùng, mà vì vậy cũng không có các telomere trừ một số trường hợp ngoại lệ. Tuy nhiên, ở những vi khuẩn có telomere, cấu trúc và chức năng của chúng lại không giống ở sinh vật nhân thực.
Đặc điểm cấu tạo của telomere:
- Có các protein đặc trưng liên kết ở phần đuôi của nhiễm sắc thể.
- Phần DNA tận cùng của nhiễm sắc thể ở dạng sợi đơn, mang trình tự lặp, cuộn lại thành dạng kẹp tóc.
Những đặc điểm nay không có ở telomere của các sinh vật nhân sơ.
Telomere trên DNA mạch thẳng được tạo thành từ trình tự lặp TTAGGG trên một mạch cùng trình tự bổ sung trên mạch còn lại (AATCCC). Một đoạn telomere là một đoạn lặp đi lặp lại của 6 cặp nitrogenous base (bazơ nitơ). Telomere ở tế bào người thông thường có khoảng 16 đoạn lặp lại như thế.
Ví dụ: Trong tế bào máu của người, độ dài của các telomere biến đổi như sau:

Mỗi lần phân bào, bình quân một người mất 30 – 200 cặp base ở các telomere của tế bào đó. Những tế bào bình thường chỉ có thể phân chia khoảng 40 – 70 lần cùng với sự ngắn dần của các telomere cho đến khi lão hóa, chết hoặc duy trì những sai sót di truyền dẫn đến ung thư. Cũng có những trường hợp telomere không bị ngắn đi. Chẳng hạn như trong mô cơ tim, do chúng không liên tục phân chia.
4. Chức năng của telomere:
Như vậy, có tthể tóm gọn lại chức năng của telomere:
– Giúp tế bào phân chia mà không làm mất gene. Nếu không có telomere, những gene cấu trúc qui định những tính trạng của cơ thể sẽ bị rút ngắn dần sau mỗi lần phân bào.
– Giữ cho các nhiễm sắc thể không bị dung hợp với nhau hoặc tự dung hợp ở những vị trí tận cùng. Khi cắt bỏ đoạn telomere ở 2 đầu của các nhiễm sắc thể, người ta phát hiện ra nhiều nhóm nhiễm sắc thể dính nhau ở đầu tận cùng, hoặc tự dính vào nhau ở 2 đầu gây nên sự khép vòng. Chức năng này có được do sự hình thành cấu trúc kẹp tóc trong telomere: phần DNA tận cùng cuộn xắn lại, giúp cho những phần DNA mạch đơn không thể kết cặp bổ sung một cách ngẫu nhiên với nhau.
– Khi đầu tận cùng của nhiễm sắc thể mất telomere, tế bào sẽ nhận diện đầu tận cùng có vẻ như bị sai hỏng (không có sự lặp lại của trình tự TTAGGG) và tiến hành các hoạt động sửa sai phần mà thực ra không có sự sai sót nào. Kết quả có thể làm dừng quá trình tự nhân đôi, thậm chí gây chết!
– Một số bệnh di truyền ngày nay được biết là do những khuyết tật của enzyme telomerase – enzyme dảm trách việc tổng hợp kéo dài đoạn telomere, duy trì tuổi thọ của các tế bào có sự tồn tại của enzyme này.
Ví dụ:
- Các dạng chính của bệnh thiếu máu bẩm sinh do tế bào gốc trong tủy xương phân chia không đủ cung cấp cho cơ thể
- Những bệnh di truyền chủ yếu trên da và phổi.
- Một vài căn bệnh gây già sớm.
5. Sự hình thành phức hệ protein – telomere ở tận cùng nhiễm sắc thể (telomere capping):
Ngoài chức năng duy trì tuổi thọ của tế bào bằng cách kéo dài thời gian tồn tại an toàn của các gen cấu trúc khi đầu mút nhiễm sắc thể “mòn” dần qua các lần phân bào, telomere còn có chức năng giữ cho các nhiễm sắc thể không bị dung hợp với nhau (fusion). Chức năng này có được là do sự hình thành phức hợp liên kết giữa các protein đặc trưng với đoạn telomere.
Các vùng giàu Guanine trên telomere chính là trung tâm giúp cho cấu trúc này được hình thành. Chúng đóng vai trò là điểm đính của các protein, được gọi là các vị trí liên kết (binding sites). Những vị trí này có cấu trúc đặc trưng cho việc đính những protein đặc hiệu phục vụ cho việc bảo vệ đầu tận cùng của telomere. Các protein hoạt động bằng cách vừa che chắn phần cuối của telomere (capping the telomere ends), vừa tham gia vào cấu hình của các cấu trúc telomere phức tạp hơn. Một cách tổng quát, các protein đóng góp vai trò vào việc bảo vệ phần tận cùng của nhiễm sắc thể khỏi sự dung hợp với các nhiễm sắc thể khá, hoặc bảo vệ chúng khỏi sự tấn công của các enzyme polimerase có hoạt tính exonuclease (phân giải acid nucleic từ đầu 3′-OH hoặc 5’OH).
– Quan niệm ban đầu về quá trình “đóng gói” telomere (telomere capping):
Nhằm đảm bảo vai trò bảo vệ các gene ở đoạn giữa nhiễm sắc thể, phần mạch kép của telomere sẽ được gắn với những protein chuyên biệt gọi là duplex DNA binding proteins (Raplp, TRF1 …) trong khi phần mạch đơn được “bao gói” bằng nhóm end-specific telomere protein (Oxytricha α/β). Như hình minh họa sau:

– Quan niệm hiện đại (được phát hiện từ năm 1999 – 2004):
Telomere có cấu trúc phức tạp hơn nhiều, và hoàn toàn đảm bảo cho việc tránh khỏi sự dung hợp của các nhiễm sắc thể tại các đầu tận cùng của chúng. Theo quan niệm này, phức hợp telomere – protein có sự tham gia cấu trúc của cả DNA và protein. Căn cứ vào sự hiện diện và vị trí của mỗi protein trong thành phần phức hợp, cơ thể sẽ “để yên” cho những đoạn DNA “lạ” mà không nhận diện chúng là những đoạn bị lỗi để rồi thực hiện sự sửa sai không cần thiết, thậm chí có thể gây sai hại nghiêm trọng.
Thành phần cơ bản của phức hệ protein trên telomere (shelterin complex hay telosome) được biểu hiện trong hình sau:

Telomere binding proteins: TRF1, TRF2, POT1, TIN2, TPP1, RAP1. Có rất nhiều enzyme tham gia vào thành phần của phức hệ shelterin (shelterin complex), trong đó có những thành phần chủ yếu:
- Nhóm bám trên các trình tự TTAGGG bằng cách nhận diện trực tiếp: TRF1, TRF2, POT1.
- Nhóm các protein cho phép tế bào phân biệt telomere với những phần DNA bị sai hỏng (không có gắn những thành phần này): TIN2, TPP1, RAP1.
5.1. Telomeric Repeat Binding Factor 1 (TRF1):
Gắn vào mạch kép bằng cách nhận diện những trình tự lặp đặc trưng của telomere.
5.2. Telomeric Repeat Binding Factor 2 (TRF2):
Gắn vào để kết nối đoạn mạch đơn và đoạn mạch kép trong telomere, làm cho telomere cong lại như hình một chiếc kẹp giấy. Chúng cũng đi kèm với TRF1 trong telosome
5.3. Protection of Telomere 1 (POT1):
Gắn vào mạch đơn của DNA và trùm lấy phần cuối của đoạn mạch kép trong telomere, đảm bảo cho phần này không bị tổn thương, hoặc bị kích thích tiến hành tổng hợp tiếp tục thành mạch kép.
5.4. Telomere Protecting Protein 1 (TPP1):
Tương tác với POT1.
5.5. TIN2:
Tương tác với TPP1
5.6. RAP1:
Tương tác với TIN2 và TRF2.
Sau khi hoàn tất quá trình gắn kết protein vào telomere, mô hình cấu trúc phức hợp có dạng như sau:

Telomere với các telosome gắn vào giữ hình dạng “cái kẹp tóc” Các protein trên bám dọc suốt telomere. Một nhóm gồm các thành phần này gọi là telosome (hay shelterin). Nhờ sự bảo vệ chặt chẽ của các protein cùng cấu trúc “cài then” mà telomere giúp các nhiễm sắc thể không có “cơ hội” gắn kết với nhau ở đầu tận cùng của chúng. Khi thực hiện thí nghiệm cắt bỏ telomere, người ta thấy các nhiễm sắc thể bị dung hợp với nhau ở đầu tận cùng. Trong thực tế, điều này sẽ gây ra những đột biến dị bội.

Sự dung hợp của các nhiễm sắc thể khi thực hiện thí nghiệm cắt bỏ telomere 6. Telomere và sự lão hóa:
Trong tế bào sinh dưỡng thông thường, cứ sau mỗi lần phân chia, telomere sẽ bị ngắn đi một đoạn khoảng 30 – 200 cặp nucleotide. Do đó, sau một khoảng thời gian nhất định khoảng 40 – 70 lần phân chia, những “chiếc nón sắt bảo vệ nhiễm sắc thể” này sẽ “mòn”, để lộ phần nhiễm sắc thể mang thông tin di truyền bên trong gây ra các sai sót nghiêm trọng đối với hệ gene, làm ngừng quá trình tự nhân đôi, hoặc thậm chí gây chết tế bào. Đây là lúc tế bào (kéo theo cơ thể) rơi vào quá trình lão hóa (senescence).
-
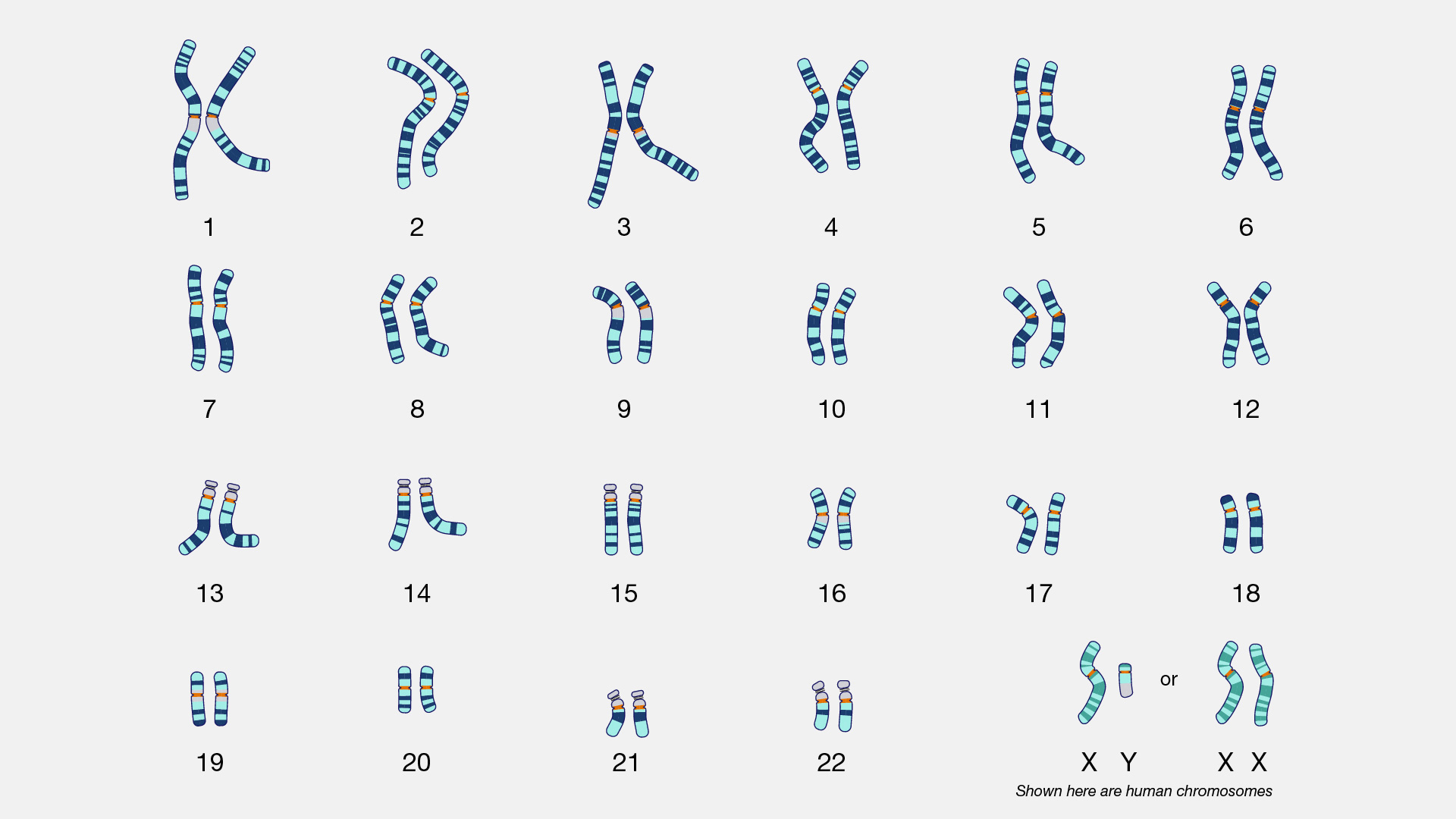
Số lượng bộ NST lưỡng bội (2n) ở một số loài
(Số lượng này liên tục được cập nhật khi có thêm thông tin mới. Mong nhận được sự giúp sức của bạn đọc)
1. Thực vật:
- Hành tây (Allium cepa): 2n = 16
- Hẹ (Allium ramosum): 2n =16
- Hẹ bông (Allium tuberosum): 2n =16
- Tỏi (Allium sativum): 2n =16
2. Động vật không xương sống:
- Tằm dâu (Bombyx mori): 2n=56
- Ruồi nhà (Rusca domestica): 2n=12
- Ruồi quả (ruồi giấm) (Drosophila melanogaster): 2n=8
3. Động vật có xương sống:
- Cá chép (Cyprinus carpio): 2n=104
- Ếch (Rana pipiens): 2n=26
- Cá sấu (Alligator mississipiensis): 2n=32
- Gà (Gallus domesticus): 2n=78
- Chuột nhắt (Mus musculus): 2n=40
- Chuột cống (Rattus norvegicus): 2n=42
- Thỏ (Oryctolagus cuniculus): 2n=44
- Ngựa (Equus calibus): 2n=64
- Bò rừng (Bos taurus): 2n=60
- Chó (Canis familiaris): 2n=78
- Mèo (Felis domesticus): 2n=38
- Khỉ rhezus (Macaca rhezus): 2n=42
- Tinh tinh (Gorila gorila): 2n=48
- Người (Homo sapiens): 2n=46
-

Cơ chế xác định giới tính
Trong chương trình THPT đề cập nhiều đến di truyền liên kết giới tính, theo đó, khảo sát sự di truyền của các tính trạng do gen nằm trên NST giới tính qui định. Việc ôn thi ĐH cũng không yêu cầu nhiều đối với việc tập trung vào cơ chế xác định giới tính. Trong nội dung học, chỉ có phần nói về cặp NST giới tính ở các loài khác nhau là xoáy vào vấn đề này. Trong chương trình thi ĐH, nội dung này chủ yếu được dùng để xác định sự di truyền ở các cá thể thuộc giới đồng giao tử hay giới dị giao tử trong một số ít trường hợp cá biệt. Do đó, việc tìm tài liệu tham khảo về vấn đề này không được dễ dàng như những vấn đề phổ biến khác của môn Sinh học.
Bài viết này không đề cập những vấn đề mang tính hàn lâm, nặng lí thuyết, mà chỉ mong tóm gọn những nội dung chủ yếu liên quan đến những đối tượng được đề cấp đến trong những bài thi cấp Bộ. Hi vọng giúp ích được cho các em học sinh và mong được trao đổi thêm cùng các đồng nghiệp.
Cơ chế xác định giới tính có nhiều kiểu khác nhau. Trong chừng mực THPT và ĐH – CĐ, chúng ta xét đến 4 cơ chế phổ biến sau:
1. Cơ chế xác định giới tính kiểu X – Y :
Ở người và các loài động vật có vú, cơ chế xác định giới tính được xác định bằng các nhiễm sắc thể X và Y. Giới cái có kiểu nhiễm sắc thể XX và giới đực có kiểu nhiễm sắc thể XY. Nhiễm sắc thể XY ở người chỉ có phần tương đồng rất nhỏ nằm ở hai đầu mút nhiễm sắc thể giúp chúng tiếp hợp với nhau trong quá trình giảm phân, phần còn lại rất lớn là không tương đồng – tức là, gen trên X không có gen tương ứng trên Y.
– Mặc dù giới tính ở người và động vật có vú khác đều được xác định theo kiểu XX – ♀️ và XY – ♂️, nhưng cơ chế này có một số điểm đặc biệt sau:
- Nhiễm sắc thể Y giữ vai trò quan trọng trong việc qui định nam tính ở người. Khi có nhiễm sắc thể Y sẽ cho ra nam giới còn nếu không có Y sẽ là nữ giới. Xem ví dụ bên dưới để thấy rõ vấn đề này.
- Trong hai nhiễm sắc thể X ở nữ giới chỉ có một nhiễm sắc thể X hoạt động còn nhiễm sắc thể kia bị bất hoạt về mặt di truyền – hầu hết các gen trên đó đều không hoạt động. Việc nhiễm sắc thể nào bất hoạt là hoàn toàn ngẫu nhiên, và được truyền lại cho một nhóm các tế bào sinh dưỡng nằm gần nhau trong quá trình nguyên phân.
Ví dụ: Năm 1990, các nhà khoa học Anh đã phát hiện ra gen SR Y (sex determining region of Y – vùng xác định giới tính trên Y) nằm ở đầu mút của nhiễm sắc thể Y qui định sự phát triển của tinh hoàn. Gen này qui định protein có chức năng điều hòa hoạt động của các gen khác tham gia vào quá trình hình thành các đặc điểm giới tính nam. Nếu không có gen này thì phôi sẽ phát triển buồng trứng và hình thành cơ thể nữ. Hiện nay người ta đã biết trên nhiễm sắc thể Y của người có 78 gen mã hóa cho khoảng 25 loại protein khác nhau (số lượng gen nhiều hơn số loại protein vì nhiều gen trong số này được lặp lại nhiều lần – đặc trưng của các gen trên nhiễm sắc thể của sinh vật nhân thực). Khoảng một nửa số lượng gen trên Y hoạt động ở tinh hoàn, một số khác cần cho sự hoạt động bình thường của tinh hoàn – tức là, hơn 50% gen trên Y được dùng để xác định giới tính nam . Chương trình giải mã bộ gen người năm 2002 phát hiện thấy trên X có 754 gen – nhiều gấp 10 lần trên Y !! [Tài liệu giáo khoa chuyên Sinh học – NXB Giáo dục]
– Ở ruồi giấm, cơ chế xác định giới tính cũng theo kiểu XX – ♀️ và XY – ♂️ như ở động vật có vú. Tuy nhiên, nhiễm sắc thể Y của ruồi giấm lại không có chức năng trong việc xác định giới tính như ở động vật có vú. Nếu phôi của ruồi giấm có 2 nhiễm sắc thể X thì sẽ phát triển thành con cái, phôi chỉ có một nhiễm sắc thể X sẽ phát triển thành con đực.
2. Cơ chế xác định giới tính kiểu X – O :
Ở một số loài châu chấu, dế và một số loài côn trùng khác, con cái có hai nhiễm sắc thể X còn con đực chỉ có một nhiễm sắc thể X. Giới tính của cá thể phụ thuộc vào việc trứng được thụ tinh bởi tinh trùng có mang nhiễm sắc thể X hay không.
3. Cơ chế xác định giới tính kiểu Z – W :
Nội dung này chủ yếu được đề cập trong chương trình bồi dưỡng học sinh giỏi hoặc ở cấp học ĐH – CĐ.
Ở chim và một số loài cá, một số loài côn trùng, con cái có nhiễm sắc thể Z và W, còn con đực có 2 nhiễm sắc thể Z. Như vậy, giới tính của cá thể phụ thuộc vào nhiễm sắc thể giới tính của trứng. Nếu trứng có nhiễm sắc thể W kết hợp với tinh trùng mang Z sẽ cho ra con cái, còn trứng mang nhiễm sắc thể Z kết hợp với tinh trùng mang Z sẽ cho ra con đực. Kiểu xác định giới tính này ngược với kiểu X – Y, và do nhiễm sắc thể giới tính trên trứng qui định.
4. Cơ chế xác định giới tính kiểu đơn bội – lưỡng bội (có ở loài động vật trinh sinh hay thực vật đơn bội)
Ở hấu hết các loài ong và kiến, tế bào không có nhiễm sắc thể giới tính riêng, và giới tính được xác định bằng mức bội thể. Nếu trứng được thụ tinh thì hợp tử 2n sẽ cho ra con cái (ong chúa hoặc ong thợ), còn nếu trứng không được thụ tinh (n) sẽ cho ra con đực.
-

Hiện tượng bù trừ liều lượng gene và sự di truyền màu lông ở thú
Trong cơ chế xác định giới tính, ở các loài động vật có vú và người, trong mỗi tế bào cơ thể cái chỉ có một nhiễm sắc thể X hoạt động, còn nhiễm sắc thể X khác thì bị bất hoạt. Tại sao có hiện tượng này? Hiện tượng này ảnh hưởng như thế nào đến sự biểu hiện của các gen di truyền liên kết trên nhiễm sắc thể này? Đến nay, hiểu biết về di truyền học đã cho phép giải thích một phần hiện tượng này.
Bà Mary Lyon, nhà di truyền học người Anh, đã đưa ra giả thuyết bù trừ liều lượng gen giải thích cho hiện tượng bất hoạt một nhiễm sắc thể giới tính ở người và các loài động vật có vú. Theo bà, sở dĩ có hiện tượng một trong 2 nhiễm sắc thể X phải bị bất hoạt là vì ở nam giới chỉ có một nhiễm sắc thể X, còn nữ giới thì lại có hai. Mặt khác, phần lớn gen trên X lại là gen qui định tính trạng thường, không liên quan gì đến giới tính – do đó mà việc chênh lệch số nhiễm sắc thể sẽ dẫn đến những tính trạng thường liên kết với nhiễm sắc thể giới tính X (không có alen tương ứng trên Y) được biểu hiện khác nhau ở hai giới. Để cho sản phẩm của các gen nằm trên X ở hai giới là như nhau, một trong hai nhiễm sắc thể X phải bị bất hoạt. Việc bất hoạt nhiễm sắc thể X nào trong cặp là hoàn toàn ngẫu nhiên.
Có một điềuđặc biệt là khi nhiễm sắc thể X nào đó trong quá trình phát triển của phôi bị bất hoạt, thì sự bất hoạt đó vẫn được di truyền cho các tế bào con trong quá trình nguyên phân. Điều này có nghĩa là một nhiễm sắc thể X có nguồn gốc từ mẹ (giả sử X1 trong cặp tương đồng X1X2) khi đã bị bất hoạt ở một tế bào phôi nào đó, thì tế bào này sẽ sinh ra các mô chứa các tế bào đều có nhiễm sắc thể X đó bị bất hoạt (trong trường hợp này là X1 trong cặp X1X2). Như vậy, có thể nói cơ thể nữ giới (hay động vật có vú giống cái) là một thể khảm gồm các vùng cơ thể có các nhiễm sắc thể X từ mẹ bị bất hoạt và vùng cơ thể khác lại có các tế bào mang nhiễm sắc thể X từ bố bị bất hoạt. Cơ chế phân tử của hiện tượng này được cho là do sự methyl hóa một số gốc cytosine (xitôzin – X) nhất định trong phân tử ADN nằm trên nhiễm sắc thể X. Nhiễm sắc thể X bị bất hoạt thường co ngắn lại, hình thành thể Barr nằm ép vào màng nhân; chất nhiễm sắc trở thành chất dị nhiễm sắc (không bắt màu).
Việc bất hoạt nhiễm sắc thể X có thể ảnh hưởng đến sự biểu hiện ra kiểu hình của các gen nằm trên nhiễm sắc thể X khi cơ thể có kiểu gen dị hợp.
Ví dụ 1: Sự di truyền màu lông ở mèo (sự xuất hiện kiểu hình mèo tam thể)
Con mèo đực thường chỉ có một màu lông đồng nhất trên cơ thể (mèo mướp – lông xám đen hoặc mèo lông vàng da cam; cả hai màu lông này có thể xen với lông màu trắng – không do gen nằm trên X qui định). Tuy nhiên, mèo cái ngoài các loại lông này còn có con có kiểu lông dạng khảm xám đen, da cam và trắng (3 màu) mà ta quen gọi là mèo tam thể. Mèo cái có lông tam thể là do nó có kiểu gen dị hợp qui định màu lông. Gen qui định màu lông này nằm trên nhiễm sắc thể X.
- Khi mèo cái có kiểu gen đồng hợp: lông có một màu (xám đen hoặc toàn vàng da cam).
- Khi mèo cái có kiểu gen dị hợp: một số vùng da mà tế bào có chứa nhiễm sắc thể X mang gen qui định màu lông đen hoạt động sẽ có lông màu xám đen. Ngược lại, sẽ có màu da cam. Điều này làm cho những vùng lông màu xám đen và da cam xen kẽ nhau.
Ví dụ 2: Ở người, gen qui định các tuyến mồ hôi nằm trên nhiễm sắc thể X. Nếu người phụ nữ có kiểu gen dị hợp thì trên da của họ có vùng có tuyến mồ hôi và vùng không có tuyến mồ hôi xen kẽ nhau – tương tự như trường hợp màu lông của mèo tam thể.
Ở ruồi giấm cũng có cơ chế bù trừ liều lượng gen nhưng khác với cơ chế bù trừ liều lượng gen ở các động vật có vú. Các gen trên nhiễm sắc thể X duy nhất ở con đực đã tăng cường mức độ hoạt động và tạo lượng sản phẩm của gen bằng với lượng sản phẩm do hai gen cùng loại ở con cái tạo ra. Do vậy, ruồi cái không có kiểu hình khảm như ở động vật có vú.
-

Bước sóng và năng lượng
Trong nội dung bài Đột biến, khi giảng về các nhân tố vật lí gây đột biến, thầy luôn gặp khó khăn khi phải ngồi giải thích lại về bước sóng cho các bạn hiểu. Thôi thì giải thích trước ở đây để tiện cho việc tra cứu của các bạn.
I. Bước sóng là gì?
Khi chúng ta quăng một hòn đá vào nước, mặt nước sẽ gợn sóng (concentric), như thế này:

Mặt nước gợn sóng khi có vật rơi xuống Trong các gợn sóng, gồm sóng nhô lên và sóng lõm xuống. Khoảng cách giữa 2 gợn sóng cùng loại liên tiếp (nhô lên hoặc lõm xuống) được gọi là bước sóng. Đây là những sóng vĩ mô và chúng ta nhìn thấy rõ các bước sóng. Có những hiện tượng sóng mà chúng ta không thấy được bằng mắt thường. Vì dụ: sóng ánh sáng, thường được đo bằng đơn vị nanometer.
Trong những trường hợp phức tạp hơn, ta có những mô tả phức tạp hơn, nhưng lại không nằm trong phạm vi thầy phải giải thích ^_^.
II. Bước sóng và năng lượng:
Chúng ta biết rằng, khi quăng một hòn đá xuống mặt hồ, hòn đá đã bị nước cản lại trước khi rơi xuống đến đáy hồ. Lực cản hòn đá do sức căng mặt ngoài của mặt nước gây nên. Theo định luật 3 Newton, sức căng bề mặt của nước tác dụng vào hòn đá, có nghĩa là hòn đá cũng tác dụng lại mặt nước một lực. Lực này sinh công, tức là truyền năng lượng cho các phân tử nước —> làm cho các phân tử nước chuyển động. Lực liên kết giữa các phân tử chất lỏng trong cả khối nước kéo nhóm phân tử nước đang chuyển động sau khi nhận năng lượng từ hòn đá kia trở về với “anh em”. Điều này gây nên sự dao động. Dao động này lan truyền đến các phân tử nước lân cận, cứ thế gây nên những dao động trên mặt hồ, lan dần đồng tâm ra khắp mặt hồ. Điều này có nghĩa là năng lượng đã lan truyền trên khắp mặt hồ.
Chúng ta nói đến kết cục khi những sóng dao động này va vào thành hồ. Năng lượng lan truyền thông qua dao động của các phân tử trên bề mặt nước đến va đập và truyền năng lượng vào thành hồ. Nhưng thành hồ rắn, nên lại theo định luật 3 Newton, “đáp trả” năng lượng về các phân tử nước, gây nên những sóng ngược chiều —> mặt nước dao động mạnh hơn.
Năng lượng của sóng càng cao (hòn đá càng bự), năng lượng sóng va đập vào thành hồ càng lớn, thì sóng ngược cũng có năng lượng lớn –> mặt nước dao động càng mạnh, có thể dẫn đến hỗn loạn. Có nghĩa là, năng lượng của sóng càng lớn thì sức công phá càng mạnh.
Bây giờ mình nói đến liên quan giữa bước sóng với năng lượng của sóng. Rõ ràng trong ví dụ trên, mỗi bước sóng của dao động sẽ mang một lượng năng lượng. Chúng ta thử so sánh giữa hai hiện tượng:
- bước sóng ngắn.
- bước sóng dài.
Trường hợp nào có năng lượng cao hơn?
Với trường hợp bước sóng ngắn, tần số (số lần) va đập cao hơn trường hợp bước sóng dài –> năng lượng truyền đến vật bị va đập cao hơn.
Ví dụ: giả sử mỗi bước sóng có năng lượng là 5kcal.
- Với bước sóng ngắn, giả sử bình quân 1 giây có 5 bước sóng va đập đến. Thì tổng năng lượng sóng truyền đến vật nhận trong 1 giây là: 5 kcal + 5 kcal + 5 kcal + 5 kcal + 5 kcal = 25 kcal.
- Với bươc sóng dài, giả sử bình quân 1 giây có 2 bước sóng va đập đến. Thì tổng năng lượng sóng truyền đến vật nhận trong 1 giây là: 5 kcal + 5 kcal = 10 kcal.
Câu hỏi là, trường hợp nào năng lượng sẽ cao hơn? Tát liên tục 20 cái trong 5 giây và 5 giây tát một lần, trường hợp nào mỏ phù nhiều hơn? ^_^
Kết luận: Với cùng mức năng lượng, ánh sáng có bước sóng càng ngắn sẽ mang năng lượng càng cao.
Điều này sẽ được thầy vận dụng trong bài giảng trên lớp về các nhân tố gây đột biến.
-
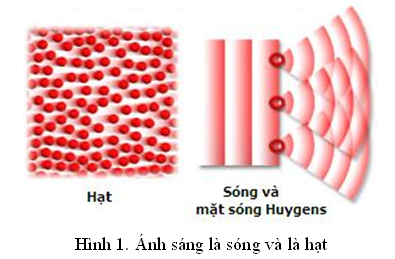
Lưỡng tính sóng – hạt của ánh sáng
Ánh sáng có lưỡng tính sóng – hạt. Điều mà học sinh lớp 12 nào ôn thi đại học cũng biết và phải biết. Nhưng môn Sinh 11 cũng có liên quan một ít đến tính chất này. Thời điểm bài QUANG HỢP được dạy trong chương trình Sinh học 11 là lúc nội dung này chưa xuất hiện trong đầu các bạn học sinh. Để bổ trợ, thầy cung cấp những hiểu biết căn bản nhất, nhằm giúp các bạn hình dung được và hiểu bài Sinh dễ dàng hơn!
I. Tính chất sóng của ánh sáng:
Trong thí nghiệm của Young, các khe sáng thứ hai, thứ ba lại trở thành những nguồn sáng thứ cấp. Chỉ những đối tượng có tính chất sóng mới làm được điều này. Để dễ hình dung, chúng ta tưởng tượng có một tấm bia, khoét một lỗ nhỏ trên đó; sau đó xả một loạt các viên đạn vào khu vực khe hở và lân cận. Giả sử rằng tấm bia đủ chắc để đạn không thể xuyên qua. Kết quả là chỉ những viên đạn có quĩ đạo xuyên ngang lỗ nhỏ mới có thể đi qua; tất cả những viên đạn nằm trong vùng cận lỗ đều bị ngăn chặn; người đứng sau tấm bia hoàn toàn an toàn nếu tránh khu vực lỗ. Những đối tượng như viên đạn được mô tả là có tính chất hạt.
Nhưng với ánh sáng, bạn sẽ không “an toàn” khi đứng sau bia, trong trường phát sáng.
II. Tính chất hạt của ánh sáng:
Trong thí nghiệm của Hertz về hiện tượng quang điện, hay đơn giản hơn, khi đi nắng chúng ta cảm thấy nóng, đều cho thấy tính chất hạt của ánh sáng – vật chất có định hình. Để dễ hiểu hơn, chúng ta xem xét một hiện tượng phổ biến khác:
Các bạn đã từng nấu nước, đúng không? Có một câu hỏi đơn giản cho hiện tượng này mà rất ít bạn học sinh thắc mắc: Tại sao nung nóng nồi mà nước cũng nóng lên? Câu trả lời là do năng lượng từ sự dao động của các phân tử trong nồi kim loại được truyền qua các phân tử nước gây gia tăng tốc độ vận chuyển của các phân tử nước. Sự gia tăng tốc độ di chuyển của các hạt trong vật thể chính là bản chất của việc gia tăng nhiệt độ.
Khi đi nắng, da chúng ta nóng lên. Điều này chứng tỏ trong ánh sáng có những hạt mang năng lượng va đập vào các tế bào trên da, cung cấp năng lượng cho da, khiến nó nóng lên. Chính điều này mình chúng rất rõ, là trong ánh sáng có các hạt đơn vị ánh sáng (photon), nên va đập vào vật thể làm chúng nóng lên.
Tức là, ánh sáng còn có tính chất hạt nữa.
Tóm lại, ánh sáng có lưỡng tính sóng – hạt.





